


宇宙のビッグバン : 全てのはじまり

空想は知識より重要である。知識には限界がある。想像力は世界を包み込む。空想は知識より重要である。知識には限界がある。想像力は世界を包み込む。
宇宙のビッグバンから地球上生物進化・人類誕生へ

宇宙ビッグバンから学ぶ 健康増進お料理レシピ
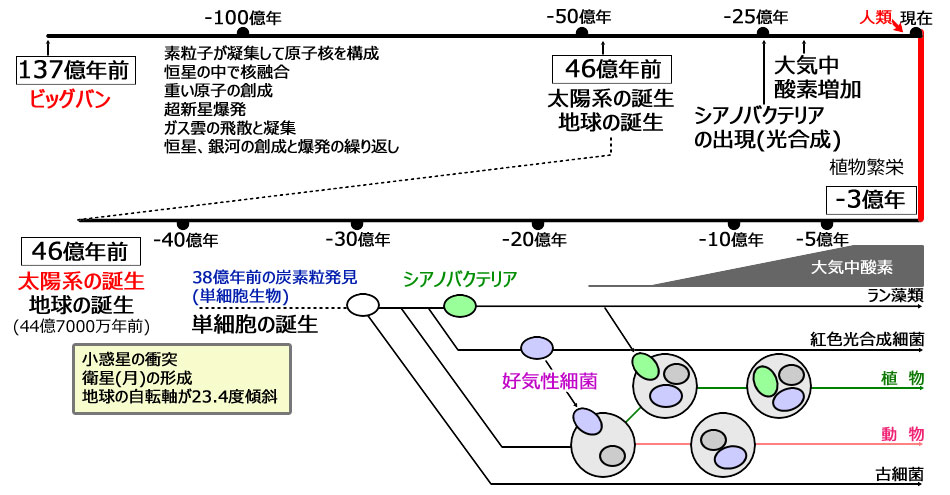
今から約137億年前、宇宙のビッグバンが起きて、私たちが日常普通に体験している時間・空間・物質が生まれたのです。ビッグバンの後約90億年の時間を経て、今から45億年前に太陽系ならびに地球を含む太陽系の惑星は誕生しました。
地球上では、RNA・蛋白質ワールドを経て、約40億年前に遺伝子情報媒体としてDNAを持つ単細胞生物の祖先が生まれ、嫌気性生物から好気性生物へ、単細胞から多細胞へ、そして動物、植物などへと進化したと考えられています。
中でも地球上生物の進化にとって革命的な出来事は、今から約20億年前に起きた大気中酸素の出現です。シアノバクテリア(珪藻類)が太陽光エネルギーを用いて、水を分解して二酸化炭素から炭化水素を作り、その”廃ガス”である酸素分子を大気中に放出し始めたのです。
一方、酸素分子を用いた高い効率のエネルギー代謝能力を持つ好気性細菌(現在のミトコンドリア)が嫌気性細胞の中に共生することによって、地球上生命体の爆発的な進化と増殖が可能になったのです。しかし、そのことは好気的エネルギー代謝の高い効率性の裏には、酸素による細胞毒性と傷害が表裏一体となって存在してきたことを意味します。
私たちの体内にあって、動脈内皮細胞は酸素化された血液に常時暴露されています。年齢と共に増える動脈硬化の患者数は、酸素分子および活性酸素による動脈内皮細胞の傷害、即ち「酸化的ストレス」に起因すると考えられます。酸化的ストレスは、動脈硬化のほかにも、癌、糖尿病、炎症等の疾患の原因・重篤化に関係しています。換言すれば、私たちの身体において酸化的ストレスは不可避の生化学現象であり、且つ疾患のリスクファクターなのです。したがって、如何に酸化的ストレスに対する防御システムを強化するか?ということが、私たちの健康増進の命題です。
活性化酸素は疾病原因、病態悪化に関係する
地球上生物の多くは、酸素を利用した好気的代謝の能力を取得して、劇的な生命進化を遂げてきた。
しかし、活性酸素による細胞障害のリスクを併せ持つようになった。
活性酸素
![]()
活性化酸素が関与すると報告されている病気 : 癌、炎症、ウィルス感染、動脈硬化、糖尿病など
 注目すべきことに、地球上で進化してきた被子植物は、自らの子孫を守るべく、ある一定の期間種子を果実の中に閉じ込めています。しかも大気中の酸素に暴露されやすい表皮と果肉には、天然の抗酸化物質を豊富に含んでいます。
注目すべきことに、地球上で進化してきた被子植物は、自らの子孫を守るべく、ある一定の期間種子を果実の中に閉じ込めています。しかも大気中の酸素に暴露されやすい表皮と果肉には、天然の抗酸化物質を豊富に含んでいます。
私たちは、地球上で独自の進化を遂げた植物から学ぶことが多いのです。即ち、健康増進のために、私たちは積極的に天然の抗酸化物質を摂取して、体内の防御システムを強化する必要があります。
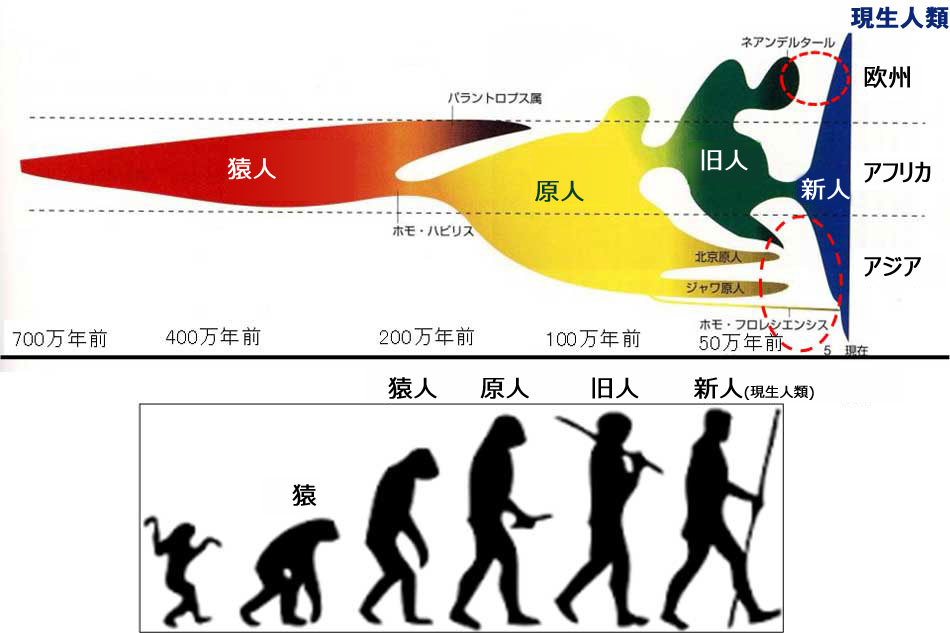
人類の大陸間移動の歴史
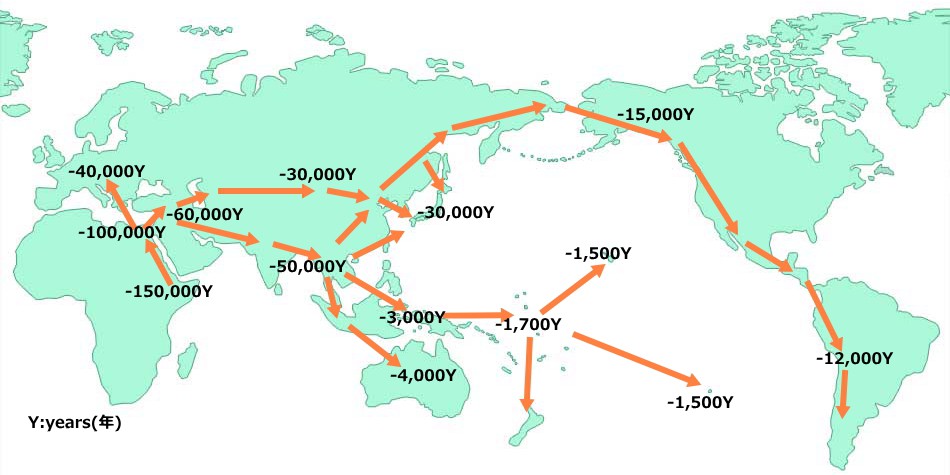
地球上生命体の進化の延長上に、私たち現生人類がいるのです。今から約20万年前に、現生人類の祖先はアフリカの東部(エチオピア付近)で生まれ、約15~10万年前にアフリカ大陸からヨーロッパ大陸、アジア大陸、アメリカ大陸、オーストラリア大陸へと大陸間移動を成し遂げたのです。その過程において、現生人類は、ネアンデルタール人やデニソニア人等とも遭遇して交配した結果、遺伝的多様性を獲得していったと考えられています。
現生人類の疾患原因と健康増進を考えるとき、地球上生命体の進化と人類の遺伝的多様性を無視することはできません。
この「宇宙ビッグバンから学ぶ健康増進お料理レシピ」の背景にある基本的哲学は、「酸化的ストレス」と「天然抗酸化物質の摂取」です。
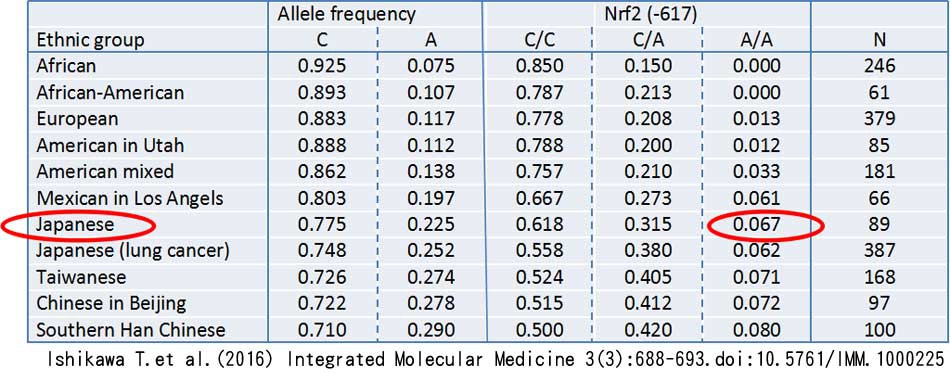
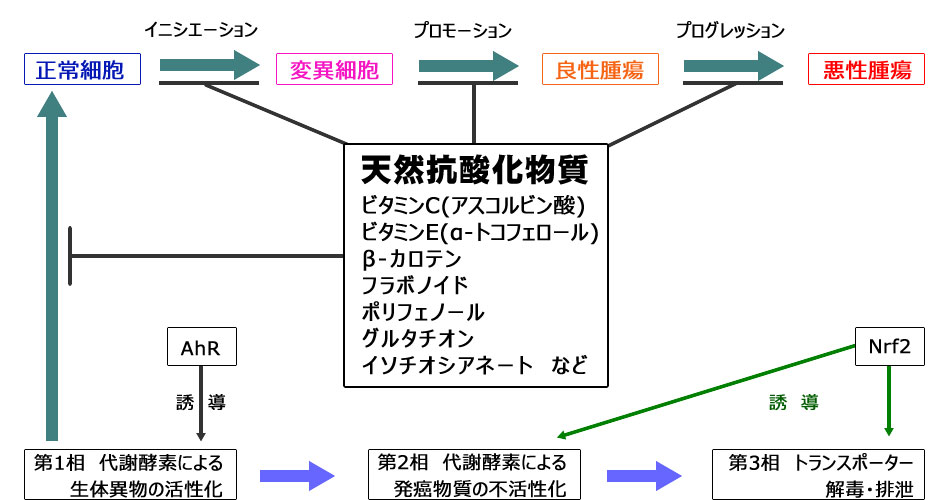
酸化的ストレスに対する抵抗能力には、私たちの体(細胞の核ゲノム)にあるNrf2という遺伝子が重要な役割を担っています。しかし、日本人の場合、6~7%の人たちは、酸化的ストレスに対して弱い遺伝子多型アレル(-617A)をホモ接合体で持っています。したがって、そのような人達は、天然抗酸化物質を積極的に摂取する必要があります。
尚、(-617A/A)ホモ接合体の割合は次の通りです。 日本人:6.7%, ヨーロッパ人:1.3%, アフリカ人:0.0%. アフリカ人は、日本人よりも酸化的ストレスに対して相対的に強いといえます。
私たち日本人は、15万年以上におよぶ大陸間移動の歴史を通して、農耕民族として飢餓に強い体と遺伝子を獲得して、それを個々の細胞の核ゲノムに持っています。つまり、少ない食事をもとに高いエネルギー源である脂肪酸を作り、飢餓に備えて脂肪酸を体内に蓄える生化学的機構をもっています。脂肪酸は糖類からも作り出されますが、飢餓時には、まず脂肪酸を使ってエネルギーを作ります。
しかし、現代社会の豊饒な食生活によって、飢餓に強い私たち日本人の体は過度の脂肪酸を作り出して腹回りなどに蓄積してしまったのです。肝臓にたまりやすい脂肪(肝脂肪)は、その1例です。毎日約1,500~2,000 kcalのエネルギーを消費している中で、使われている燃料の比は:炭水化物60%、脂肪酸25%、蛋白質15%です。脱・炭水化物生活を継続することによって糖類の摂取量を減らして、体内に蓄積した脂肪酸を主たるエネルギー源をして利用させることができます。すると体脂肪率が低下してきます。飢餓の際には、蛋白質をエネルギー源にする際には、オートファジー機構が働いて蛋白質を分解します。そして、できたアミノ酸をエネルギーに変換して、私達の体は飢餓時に生命を維持することができます。
一方、毎日の食生活に野菜や果物は欠かせません。ビタミン類など、私たちの体内で作り出せない成分を、食物から摂取する必要があります。野菜の鮮やかな色は、体のために必要な成分がギュッと詰まった証拠です。大きく赤・橙・黄・紫・緑・褐色・白の7色で分けられており、特に野菜を摂るときはこの色をバランス良く摂取するのがいいのです。それは人類が地球上に現れて以来、命を賭けた試行錯誤の過程で獲得した生活の知恵なのです。
赤色の野菜
赤色の野菜の代表はトマト。トマトに含まれる赤色の色素「リコピン」は強い抗酸化作用があることで知られています。一方、赤色の仲間の赤唐辛子には「カプサイシン」が豊富に含まれていて、私達の体内の新陳代謝を促進します。
橙色の野菜
ニンジンやかぼちゃなど、橙色の野菜からは「プロビタミンA」を摂取できます。なかでもα-カロテンやβ-カロテンが有名で、これらの栄養素が体内でビタミンAに変換されます。ビタミンAは天然の抗酸化物質であり、肌の健康維持等を助けてくれます。
黄色の野菜
黄色の野菜には、トウモロコシや黄色のパプリカ、大豆などが含まれます。トウモロコシなどに含まれる「ルテイン」はポリフェノールの一種で、加齢による視力の低下を予防すると考えられています。
紫色の野菜
紫キャベツや紫玉ネギ、ナスなどの紫色は「アントシアニン」というポリフェノールの一種によるものです。蒲萄の果皮にもその物質が豊富に含まれています。アントシアニンは強力な抗酸化作用を持っています。
緑色の野菜
ブロッコリーやホウレンソウといった葉物野菜や、多種類の緑色の野菜がこのグループに属します。緑色は「クロロフィル」によるもので、植物の光合成には不可欠の物質です。クロロフィルの中心にはマグネシウム・イオンがあり、それが私たちの体内の代謝にとって重要な成分です。
褐色の野菜
ジャガイモやゴボウなど、根菜は褐色のグループです。これら褐色の野菜に含まれるのは、ポリフェノールの一種である「クロロゲン酸」です。この色素成分には日焼けによるメラニンの生成を抑えたり、糖の吸収を遅らせる働きがあるといわれています。
白色の野菜
大根やカブなど、白色の野菜に含まれる「硫化アリル」は、疲労回復を助けるビタミンB1の吸収を促す働きがあります。また血糖値の上昇を抑制する効果も期待されています。
宇宙ビッグバンから学ぶ 健康増進お料理レシピ

















あとがき
「食べる」ことは、生命の維持にとって必要不可欠な行為です。私達は毎日無意識のうちに野菜や果物、米、パン、麺、魚、肉などを食べています。しかし、それらは全て地球上の生物(植物・動物・微生物)に由来するのです。
137億年前に起きた宇宙のビッグバンの前には、時間も空間も物質もなかったのです。唯一あったのは一点に集約された膨大なエネルギーだったのです。そこから「ビッグバン」によって、今のような宇宙が形作られてきたのです。地球が太陽系に生まれたのは今から44億7000万年前と考えられています。そして、約40億年前には原始生命体が生まれ、38億年前には単細胞生物が出現したと報告されています。
私たち、現生人類が出現したのは約20万年前の短時間であり、その1万倍以上の時間スケールで地球上生物の進化・淘汰・絶滅という壮大なドラマが繰り広げられてきました。その最後の一瞬の果実を、私たちが享受しているのです。
これまで約40億年の歴史:地球上生物の進化
- 化学進化(生命体を構成する物質の創成)
- リボ核酸(RNA)とたんぱく質の複合体
- 原始生命体の誕生&ウイルス祖先の出現
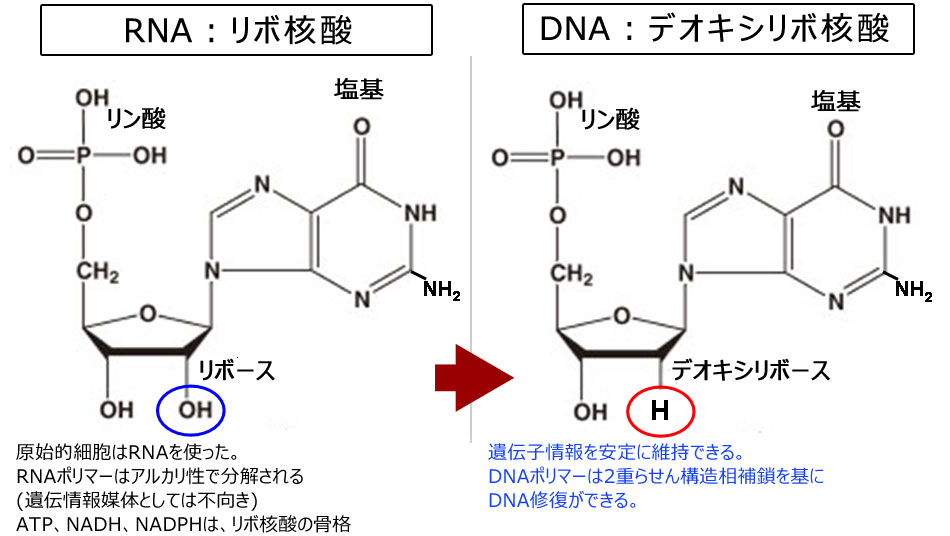
- 遺伝情報媒体の変化(RNA⇒DNA)
- 単細胞生物の誕生(無性増殖)
- 嫌気性細胞への葉緑体およびミトコンドリアの共生
- 原核細胞⇒真核細胞
- 単細胞⇒多細胞
- 進化を加速させた活性酸素・紫外線・放射線
- 遺伝子変異による進化の多様性
- 遺伝子修復機能と子孫への遺伝情報伝達
- 雌雄交配(有性生殖)による増殖・遺伝子多様化
- 水中生物⇒陸上生物
- 環境への適用(重力・紫外線・活性酸素に対応する対応)
原始生命体の誕生とウイルス祖先の出現
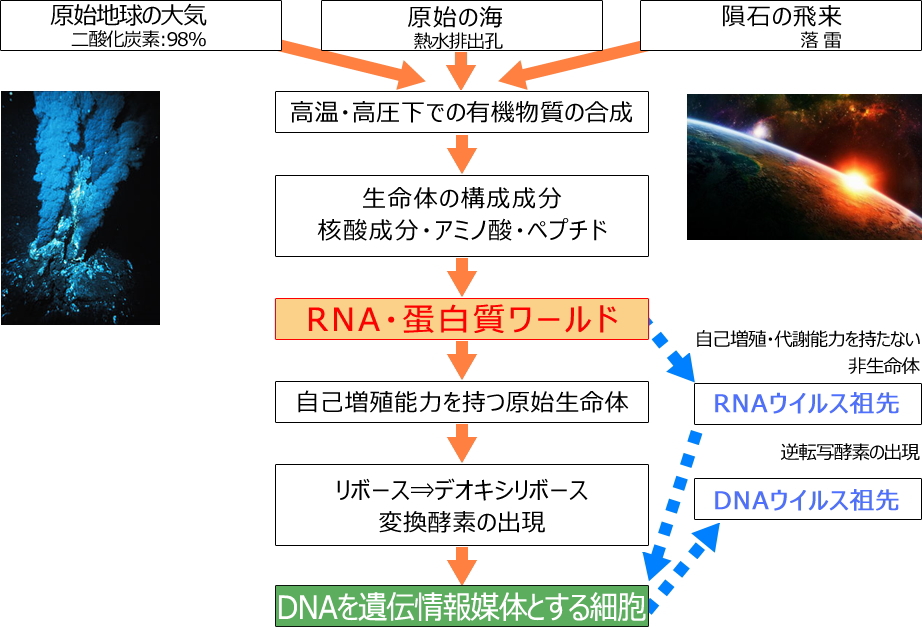
遺伝子情報媒体:RNAからDNAへ分子進化
植物の出現 光合成によって大気中に酸素を放出

猿から現生人類への進化
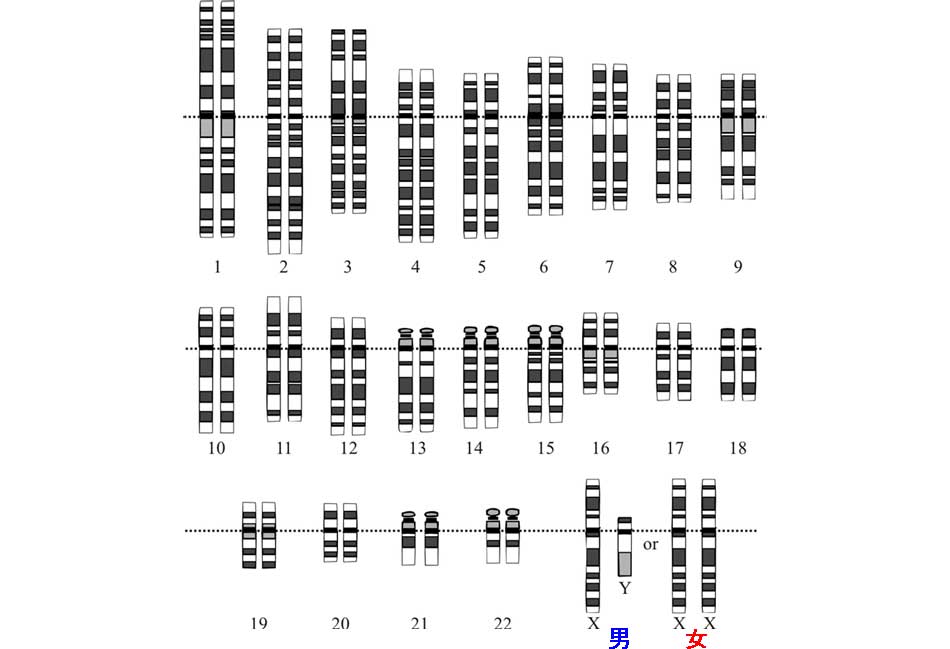
人類の染色体:遺伝情報を保持する複合体
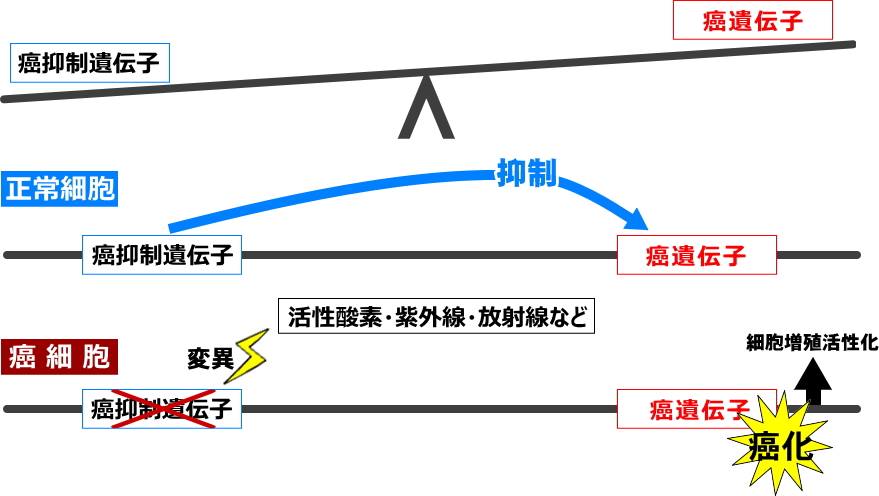
癌抑制遺伝子vs.癌遺伝子
天然成分摂取による癌の予防
地球上生物の進化の過程の中で、細胞の増殖を加速する癌遺伝子がレトロウイルスなどによって組み込まれてきました。その結果、私たちの染色体にあるゲノムDNAの中には既に、細胞の増殖を加速する癌遺伝子と、その活動を抑制する癌抑制遺伝子とが共存しています。もし活性酸素、ラジカル、放射線、紫外線などによって癌抑制遺伝子に変異が起きてしまうと、発癌プロセスに”ON”スイッチが入ってしまいます。そして正常細胞から悪性腫瘍に変化するには多段階を経て進行します。
一方、私たちが摂取する食べ物の中には天然抗酸化物質が含まれており、それらが体内で癌化のプロセスにブレーキをかけています。
料理に含まれる天然成分:健康増進の科学
食べ物の消化・吸収・解毒・代謝
地球上ほとんどの生物は、成長や増殖、生命維持などに必要な成分を「食べ物」から摂取しています。私たちの体において、食べ物は、消化・吸収の過程を経て、生命の維持や運動に必要なエネルギーの産出、または生体成分へと転換されて利用されています。
味覚は、食べ物の安全性を評価するうえで重要です。有毒物質が食べ物に含まれていた場合、私たちの体は嘔吐および下痢によって、有毒物質が体内血液循環に入るのを防ぎます。咀嚼・嚥下に続く消化行動のあと、食べ物中の栄養物質は消化管(特に小腸)で吸収され、肝臓で解毒・代謝を受けます。そして心臓の血液ポンプ機能によって様々な組織・細胞に送られるのです。
栄養物質を体内へ 毒性物質を体外へ

味覚の受容体
味を感知する受容体(レセプター)には、沢山種類のあることが判ってきました。甘味、旨味、苦味に対する受容体はいずれも、細胞膜を貫通する領域を7つ持った「Gタンパク質共役受容体(G protein-coupled receptor, GPCR)」という受容体のグループに属しています。この甘味、旨味、苦味の味覚受容体タンパク質はその構造的特徴から、大きな細胞外領域を持つものと、この 領域を持たないものの二種類に大別でき、前者をT1Rファミリー、後者をT2Rファミリーと呼びます。
T1Rファミリーには、T1R1〜T1R3までの三種類があって、これらのうち、T1R1とT1R3の組み合わせで旨味受容体を、T1R2とT1R3の組み合わせで甘味受容体を構成していることがわかりました。苦味受容体であるT2Rファミリーには多くの種類があり、ヒトでは26種類が見つかっています。
おいしさの科学
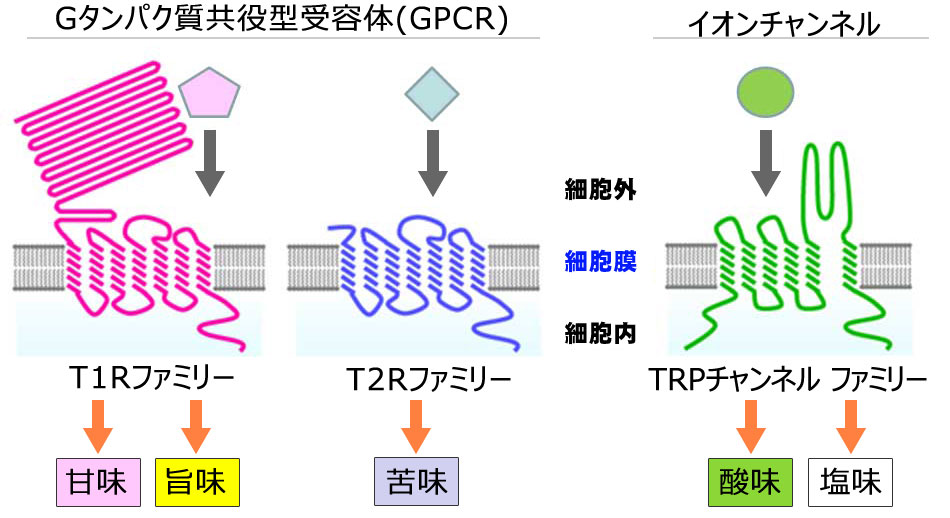
一方、酸味や塩味に対する受容体はイオンチャネル型の分子だと考えられています。TRPチャネル(transient receptor potential channel、一過性受容体電位型チャネル)と呼ばれ、これまでに温度や辛みなどの受容に関わることが知られていたチャネル分子の仲間であろうと考えられています。Gタンパク質共役型受容体が味物質と結合してGタンパク質を活性化するのとは対照的に、イオンチャネル型受容体は、細胞外のH+(酸味)やNa+(塩味)などのイオンによって開口し、陽イオンを透過させるイオンチャネルとして働くことにより、味細胞を脱分極させて、神経にシグナルを伝えるのです。
味覚受容体と味物質
| 味 | 受容体 | 味物質 |
| 甘味 | T1R2+T1R3 |
糖類(ショ糖、ブドウ糖、果糖、麦芽糖) 人工甘味料(サッカリン、アスパルテーム、アセスルファムK、シクラメート) 甘味タンパク質(モネリン、クルクリン) |
| 旨味 | T1R1+T1R3 |
アミノ酸(L-グルタミン酸)、核酸(イノシン酸) |
| 苦味 | T2R5 |
シクロヘキシミド |
| T2R4,8,44 |
デナトニウム |
|
| T2R16 |
サリシン |
|
| T2R38 |
フェニルチオカルバミド(PTC) |
|
| T2R43,44 |
サッカリン |
|
| 不明 |
キニーネ、ストリキニーネ、アトロピン、カフェイン |
|
| 酸味 | PKD2L1+PKD1L3 |
酸(クエン酸、酒石酸、酢酸) |
| 塩味 | ENaC TRPV1 |
塩化ナトリウム |
味シグナルの伝わる仕組み
味の感知は舌上皮に存在する味蕾と呼ばれる組織中にある特殊な細胞で行われます。味蕾の味覚受容体で感知された味シグナルは、舌の前半部にある鼓索神経、あるいは舌の奥にある舌咽神経を伝わって延髄孤束核に伝えられます。孤束核では、神経の乗り換えが起こり、味覚情報は視床の味覚野へ投射され、大脳皮質味覚野では、味の種類、強さが識別されます。したがって、料理の味を楽しむことは、大脳皮質を活性化するうえでも重要なのです。「臭い」と並んで、「味」は大脳皮質の衰えを抑制するうえでも重要なファクターです。
謝 辞
亡き愛妻・直子に この「宇宙ビッグバンから学ぶ健康増進お料理レシピ」を捧げます。
32年間の結婚生活の間、妻・直子がずっと私を精神的に支えてくれましたが、2014年7月に卵巣癌の末期(stage 4)と診断されて、神奈川県立がんセンターに入院して治療を受けました。しかし、12月10日未明3時過ぎに永眠しました。約5か月間彼女を看病し続け、最期の6時間は彼女と並んで寝て、彼女の肩を抱きながら「世界一好きや、愛してるんや」と何度も言い続けました。亡くなった時、彼女の左目には涙が光っていました。
彼女の葬儀が終わってから2か月間、私は精神的な支えを失いダウンしそうにもなりました。自分一人で料理を作って、食べるという淋しく暗い日々が続きました。しかしある日、私の作った料理を写真に撮影して、そのレシピをFaceBookにアップロードしたら、沢山の方々から激励の言葉と「いいね」を頂きました。とても有難かったです。それがきっかけとなって、再び生きる力が湧いてきて、創作料理に精を出すことになったのです。そうして過去1年半の間に私が作った創作料理をまとめたのが、この料理レシピ集です。
1982年10月、私はドイツDüsseldorf大学医学部の生理学研究所でHelmut Sies教授とともに酸化的ストレスの研究を始めました。Sies教授は、Oxidative Stressという新分野を開拓して世界をリードしてきました。彼の影響をうけて、私は酸化的ストレスと疾患という研究テーマに取り組むようになりました。したがって、この「宇宙ビッグバンから学ぶ健康増進お料理レシピ」の根底には、「酸化的ストレス」の考え方が流れています。
Helmut Sies教授と私は、ひとまわり違いの午(うま)年生まれで、彼は私達夫婦をいつも気にかけてくれました。2014年3月にSies教授夫妻を訪問して、彼らの家に泊めてもらいました。その時に撮影したのが次の写真です。それが、4人そろった最後の思い出の写真になりました。Helmut Sies教授とその妻Nancyに心より感謝いたします。


略 歴

| 氏 名 | 石川 智久 (いしかわ としひさ) |
| 学 位 | 理学博士 |
| 生年月日 | 1954 年9 月4 日 |
| 職 業 |
NPO 法人 地方再興・個別化医療支援 理事長 大阪医科大学 大学院医学研究科 客員教授 |
学歴および職歴
| 1967年3月 | 西条市立神戸小学校 卒業 |
| 1970年3月 | 西条市立南中学校 卒業 |
| 1973年3月 | 愛媛県立西条高校・理数科 卒業 |
| 1973年4月 | 北海道大学・理類 入学 |
| 1977年3月 | 北海道大学・理学部・化学科 卒業 |
| 1982年3月 | 北海道大学大学院・理学研究科博士課程 修了 理学博士取得 |
| 1982年4~6月 | 学術振興会・奨励研究員(北海道大学・応用電気研究所・生体物理部門) |
| 1982年6~9月 | ドイツ学術交流会 (DAAD) 奨学生として渡独、4 か月間ドイツ語研修 |
|
1982年10月 ~1987年4月 |
ドイツDüsseldorf 大学医学部 生理化学研究所 Helmut Sies 教授の研究室にて博士研究員として基礎医学研究に従事 |
| 1987年4月 | 大阪大学医学部 生化学教室 (谷口直之教授) の助手に採用 |
| 1989年10月 |
ドイツ癌研究所 (Heidelberg) 腫瘍生化学部門 研究員 重点領域研究 (SFB) プロジェクトリーダーに就任 |
| 1991年11月 |
テキサス大学 M.D.アンダーソン癌センター小児癌研究部門 助教授に就任 テキサス大学 医科学研究センター大学院 助教授を兼任 |
| 1995年12月 | ファイザー製薬中央研究所 生物科学研究室 主任研究員を経て室長に就任 |
| 1998年9月 | 同研究所 学術交流推進室 室長に就任 |
| 1999年12月 | ファイザー製薬東京本社PHA 研究技術開発担当部長に就任 |
| 2000年6月 | 東京工業大学 大学院生命理工学研究科 生体分子機能工学専攻・教授に就任 |
| 2002年4月 | 大学発バイオベンチャー 株式会社メディシナル・ゲノミクスを設立 (現在名:株式会社ジェノメンブレン) |
|
2009年5月 ~2014年3月 |
理化学研究所・オミックス基盤研究領域・客員主管研究員を経て横浜研究所・主幹、上級研究員 理化学研究所 創薬・医療技術基盤研究プログラム・テーマリーダー |
| 2012年4月~ | 横浜市立大学 大学院医学研究科・客員教授 |
| 2014年3月~ | 大阪医科大学 大学院医学研究科 客員教授 |
| 2014年4月~ | NPO 法人 地域再興・個別化医療支援 理事長 個別化医療研究所・所長 |
受 賞 歴
| Deutscher Akademischer Austauschdienst (DAAD)奨学金(1982-1983) |
| International Life Sciences Institute (米国.) Achievement Award 受賞 (1993) |
| 第1回バイオビジネスコンペJAPAN 優秀賞受賞 (2001) |
| 日本薬物動態学会 (JSSX) 学会賞受賞 (2005) |
| JSSX Fellow 称号 (2008) |
| Drug Metabolism & Pharmacokinetics 最優秀論文賞(2008) |
国際活動
| HUGO ヒトABC トランスポーター遺伝子命名法国際委員会メンバー |
| FDA Critical Path Transporter Working Group 運営員会メンバー |
| International Transporter Consortium 組織委員会メンバー |
| ILSI Health Environment Science Institute(HESI) Emerging Issue Committee の委員会メンバー |
| NIH Pharmacogenetics Research Network (PGRN) Affiliate member |
| FEBS ATP-Binding Cassette (ABC) Proteins: Scientific Advisory Board メンバー |
| 2015 Gordon Research Conference “Multi-Drug Efflux Systems”の大会長 |